Большеухая лисица
Otocyon megalotis (Desmarest, 1822)

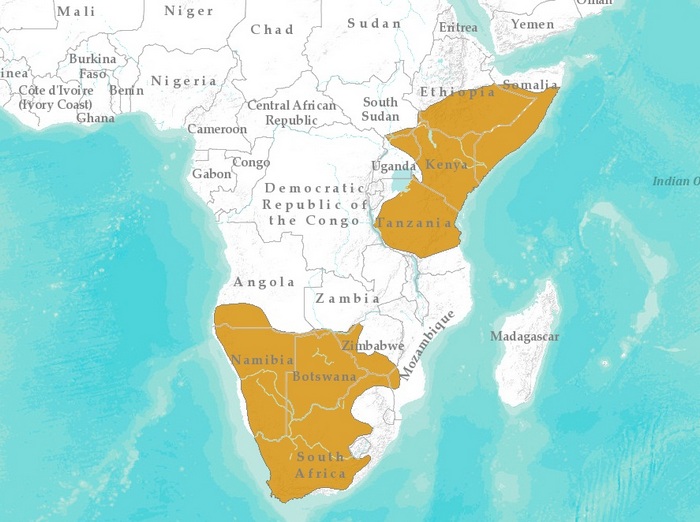
Ареал: разделен на 2 аллопатрические популяции; северная (O. m. virgatus) - Восточная Африка от Эфиопии, Сомали и Южного Судана через Кению до Танзании; южная (O. m. megalotis) - Южная Африка от южной Замбии и южной Анголы до ЮАР, на восток до Мозамбика, Ботсваны и Зимбабве, на запад до Атлантического океана.
Страны ареала: Ангола, Ботсвана, Зимбабве, Кения, Мозамбик, Намибия, Сомали, Судан, Танзания, Уганда, Эфиопия, ЮАР.
Маленький представитель псовых с тонкими ногами, длинным густым хвостом и заметно большими ушами. Самцы (4,1 кг) тяжелее самок (3,9 кг) (в среднем для обоих полов 3,9 кг), хотя в Ботсване самки весят незначительно больше, чем самцы.

Голова, спина и верхние части ног серые. Морда черная сверху и белая по бокам. Грудь и нижняя часть тела варьируются от бледно- до медово-желтой. Уши белые внутри. Задняя сторона ушей, передняя часть морды, маска на лице, фронтальная часть передних ног и нижние части задних ног, срединная дорсальная область хвоста черные. Белесоватая полоса простирается от лба до нижних и верхних 3/4 переднего края ушей. У некоторых особей по спине проходит широкая темная срединная полоса. От бежевого до медового цвета мех покрывает нижнюю челюсть от конца морды и простирается через горло, грудь на нижние части тела. Окраска более старых особей бледнее. Мех на теле и хвосте плотный и мягкий, на верхних частях волосы черные у основания с белыми кончиками, придающими шерсти седоватый или серый вид. Бока выглядят более желтоватыми. Подшерсток на верхней части тела имеет длину около 30 мм, а остевые волосы плотного меха - около 55 мм, с разбросанными тактильными волосками (до 65 мм).
Зубов 46-50, что является наибольшим числом для любого не сумчатого наземного млекопитающего.
У самок 4-6 сосков.
Диплоидное число хромосом 2n = 72.
Длина головы и тела (полы объединены) 46-66 см, длина хвоста 23-34 см, высота плеча 30-40 см; высота уха 11,3-13,5 мм; масса 3,0-5,3 кг.
Сходный вид южноафриканская лисица (Vulpes chama) - несколько меньше; имеет серебристо-серые верхние части тела, нижние от белого до бледного палевого; голова и задняя сторона ушей красновато-палевые; нет черного на спине; хвост более густой, только кончик черный.

Дизъюнктивный ареал в засушливых и полузасушливых районах восточной и южной Африки в двух дискретных популяциях (представляющих отдельные подвиды), разделенных примерно на 1000 км. Два ареала, вероятно, были связаны во время плейстоцена. Такое дизъюнктивное распространение аналогично ареалу земляного волка (Proteles cristatus) и чепрачного шакала (Canis mesomelas). Расширение ареала в южной Африке в последние годы связано с изменением осадков.
Вид распространен в природоохранных районах в Южной и Восточной Африке, редок в засушливых районах и на фермах в Южной Африке, где их иногда преследуют. В пределах ограниченной среды обитания количество может колебаться от многочисленного до редкого в зависимости от количества осадков, наличия корма, стадии размножения и болезней.
На юго-западе Калахари численность может меняться со временем: регулярные подсчеты вдоль 21-км участка сухого русла реки площадью около 10 км² составили 7-140 особей, т. е. 0,7-14 на км². В провинции Лимпопо, Южная Африка, плотность 5,7 лисиц на км², а в близлежащем заповеднике Машату, Ботсвана, 9,2 лисиц на км² в сезон размножения и 2,3 лисицы на км² в другое время. В заповеднике Tussen-die-Riviere, провинция Фри-Стейт, Южная Африка, плотность варьировалась от 0,3-0,5 лисиц на км² в течение трехлетнего периода, тогда как на двух фермах в центральном Кару, Северо-Капская провинция, плотность составила 1,1-2,0 лисиц на км². В Серенгети зафиксирована плотность 0,3-1,0 лисиц на км².

Большеухие лисицы адаптированы к преимущественно насекомоядному рациону. Огромные уши, используемые для обнаружения насекомых, являются наиболее заметной морфологической адаптацией и также имеют функцию терморегуляции. Кормление насекомыми повлияло на количество и форму зубов лисиц.
Ареалы распространения большеухих лисиц и термитов Hodotermes и Microhodotermes перекрываются на 95%. Термиты (Hodotermes mossambicus) составляют до 80-90% рациона. В районах без Hodotermes лисицы употребляют другие виды термитов; Odontotermes также составляет более 90% рациона в некоторых частях Кении. Другие потребляемые беспозвоночные включают муравьев (Hymenoptera), жуков (Coleoptera), сверчков и кузнечиков (Orthoptera), многоножек (Myriapoda), бабочек и их личиночные формы (Lepidoptera), скорпионов (Scorpionida) и фаланг (Solifugae). Также в рацион входят птицы, мелкие млекопитающие и рептилии. Случайно съедают траву во время кормления насекомыми. Ягоды, семена и дикие фрукты потребляются уже специально. Когда ягоды осыпаются большеухие лисицы из логова прямиком следуют к известному им месту и едят плоды. Добыча птиц и поедание падали представляет собой случайную возможность, а не предпочтение. Отказывались есть термитов Trinervitermes trinervoides, экспериментально смешанных с маслом и добавляемых к их рационам, видимо, из-за того, что не терпят химических защитных выделений солдат-термитов.
Пропорции разных таксонов в рационе меняются сезонно. В Серенгети навозные жуки являются основным источником пищи в сезон дождей, когда активность термитов снижается. Когда мало и тех и других, часто выкапываются из земли личинки жуков.

Термиты и навозные жуки более распространены в районах, заселенных группами семей большеухой лисицы, а локальные различия плотности H. mossambicus обратно пропорциональны размеру занимаемой лисицами территории. Плотность выходов из гнезд Hodotermes положительно связана с различными демографическими и репродуктивными переменными, такими как размер помета и коэффициент размножающихся самок. Хотя потребность животных в воде может быть удовлетворены высоким содержанием влаги из добываемых насекомых или ягод летом в южной Африке, вода является критическим ресурсом в период лактации. Однако другие источники указывают, что нет наблюдений, чтобы лисицы пили из открытых источников воды.
Не отмечено, чтобы большеухие лисицы припрятывали излишки пищи или охотились больше, чем могут съесть. В зоопарках они обычно просто оставляют недоеденное мясо.

Техника кормления зависит от типа жертвы, но пища часто обнаруживается медленным передвижением лисицы, опустившей нос к земле и поднявшей уши вперед. Местоположение добычи определяется главным образом по звуку; зрение и обоняние играют меньшую роль. Изменения в ежедневной и сезонной доступности H. mossambicus напрямую влияют на модели активности лисицы. В восточной Африке преобладает ночное кормление. В южной Африке ночное кормление летом постепенно меняется на почти исключительно дневное зимой, отражая изменения активности H. mossambicus. Днем пики активности кормления соответствуют высоте активности насекомых. Насыщение и скорость кормления выше при кормлении термитами, чем более рассредоточенными насекомыми (например, личинками жуков или кузнечиками).
В Серенгети, покинув вечером логово, группы часто патрулируют известные им места обитания Hodotermes на своей территории. При кормлении на термитных участках члены группы кормятся близко друг к другу, но при кормлении жуками, личинками жуков или кузнечиками могут отделяться до 200 м друг от друга. Члены группы общаются между собой в богатых кормом участках при помощи низкого свиста.
В отношении хищничества домашнего скота регистраций нет. Однако в Южной Африке большеухих лисиц иногда ошибочно принимают за вредных хищников, когда те кормятся личинками мух на трупах ягнят.

Чаще всего встречаются на лугах с короткой травой (высота травы 100-250 мм) и в саваннах в засушливых и полузасушливых районах, но при угрозе прячутся в высокой траве или густых кустарниках. Во время сильных ветров и при низких температурах укрываются в растительности или в самостоятельно вырытых логовах. Лисицы переделывают существующие отверстия и используют небольшие ямки для укрытия от солнца в середине дня. Предпочитают голую землю или траву, укороченную выпасом копытных или сжиганием, в Южной Африке часто отдыхают под деревьями рода Acacia.
Активность может быть дневной или ночной, в зависимости от сезона и обстоятельств. Дневная активность тесно связана с активностью насекомых, особенно термитов. Лисицы из сухого русла реки Носсоб (национальный парк Калахари Гемсбок, Южная Африка) тратили 70-90% своего времени на кормление, но их активность варьировалась в течение года. Зимой группы лисиц в русле были активны днем, все особи, наблюдавшиеся ночью, лежали и спали. Во время южноафриканских летних месяцев декабря и января цикл активности изменился.

Зарегистрированные размеры домашнего участка варьируются от 0,3 до 3,5 км². Домашние участки групп показывают существенное или малое наложение одного участка на другой. Предпочитают сгруппированную добычу (колонии термитов); это приводит к более высокой плотности и меньшим домашним участкам при кормлении термитами (15-19 лисиц в пределах 0,5-5,3 км²), чем при потреблении другой добычи. Домашние участки меньше во время южноафриканской зимы, когда термиты составляют большую часть рациона, чем летом.
Размер группы варьируется в зависимости от времени года и колеблется от 2 до 15 лисиц. Отец охраняет логово и щенков, в то время как мать кормится для вырабатывания молока. Семейные группы кормятся вместе с декабря по июль, после чего распадаются. Наиболее часто встречаются группы по 2 особи. Большие группы взрослых состоят из родителей и их взрослых потомков. Известные пары и группы не были обнаружены в следующем сезоне размножения в данной области. Это означает, что большеухие лисицы не используют одну и ту же территорию их года в год. Вне сезона размножения, когда трава становится слишком высокой лисицы покидают свои участки.

На юге Африки живут в моногамных парах с детенышами, в то время как в восточной Африке живут стабильные семейные группы, состоящие из самца и до 3 тесно связанных самок с детенышами. Размножение происходит сезонно и локально, чтобы роды совпадали с дождями и пиком плотности насекомых. Сезон размножения с июня по сентябрь в Серенгети, в январе в Уганде. Размножение может происходить круглый год в некоторых частях Восточной Африки. В Калахари образование пар, происходит в июле и августе, о чем свидетельствует поведение маркировки территории. В Восточной Африке щенки рождаются с конца августа до конца октября, в Калахари с сентября по ноябрь. Рождения в Ботсване происходят с октября по декабрь.
Спаривание длится несколько дней (до 10 совокуплений в день), с копулятивным склещиванием, продолжающимся около 4 мин., за которым следует своеобразная пост-копулятивная игра.

Размножающаяся пара выкапывает логово или переделывает брошенное логово других млекопитающих (например, долгоногов Pedetes spp., трубкозубов, и даже курганы термитов и норы бородавочников Phacochoerus spp.). Логова могут иметь несколько входов, камер и туннелей длиной до 3 м и использоваться для защиты от хищников и стихии (например, от затопления, экстремальных температур), особенно новорожденных щенков. Детенышей иногда перемещают между логовами, а в Серенгети лисицы используют «кормовые логова» для защиты детёнышей в разных частях участка. За логовами тщательно ухаживают в течение года, часто разные поколения. Натальные логова могут быть сгруппированы: на юго-западе Калахари в 1976 году было обнаружено шесть логов на отрезке 0,5 км² русла, и каждое из них было занята взрослой парой и 2-3 детенышами (всего 16). Рядом были еще два логова.

Помет 1 раз в год, с рождениями с октября по декабрь, после беременности 60-75 дней. Размер помета колеблется от 1 до 6, в Серенгети в среднем - 2,56. Новорожденные весят от 99-142 г. Маленьких щенков выращивают внутри логова, позже снаружи. Впервые щенки ненадолго появляются из логова когда им 8-12 дней.
Самец проводит больше времени с детенышами, чем самка. Он ухаживает за ними, играет, охраняет и защищает их от хищников. Материнский вклад в выращивание потомства во время лактации высок, но из-за насекомоядного рациона, она не может в обычном смысле ухаживать за щенками. Однако высокий уровень родительской опеки самца позволяет самкам максимизировать свое время кормления, которое ограничено небольшими, рассредоточенными объектами питания. Несоответствие по уходу между полами родителей становится менее выраженным после прекращения кормления молоком (в 10-15 недель), которое на юго-западе Калахари происходит после первых дождей и последующего изобилия насекомых.

Молодые детеныши инициируются к кормлению самцом, который их водит за собой, а в Серенгети родители облегчают доступ маленьким и уязвимым детёнышам к различным группам H. mossambicus, регулярно направляя детенышей из натального логова в «ночные кормовые логова».
Ядро семейной группы продолжает существовать до следующего июня, когда молодые уходят с участка, а пара остается вместе на всю жизнь. Большинство молодых рассеивается в возрасте ок. 5-6 мес., но половая зрелость наступает несколько позже, в 8-9 мес. Некоторые молодые самки остаются со своей семейной группой для размножения.
В основном насекомоядные и не приносят добытых насекомых в логово.
Вместо этого молодым взрослые приносят некоторую добычу из позвоночных. В целом, щенки сильно зависят от молока.

Большеухие лисицы общественные животные. Кормятся группами, редко далее 200 м друг от друга и обычно находятся на расстоянии менее 30 м на открытой местности. Отдыхают вместе и часто занимаются груммингом друг друга. Коммунальное кормление семейными группами - это стратегия против хищников и метод использования насекомых.
Собираться вместе тесной группой - обычное явление у щенков и взрослых, взаимное ухаживание за шерстью между молодыми и взрослыми происходит в течение всего года. Во время таких тесных собраний подбородок одной лисицы обычно опирается на крестец другой. Во время взаимного ухаживания основное внимание уделяется лицу. Взаимное ухаживание за шерстью - преобладающий социальный контакт между взрослыми. Члены группы могут собираться в логовах ночью или ранним утром, чтобы не замерзнуть, или чтобы избежать зноя. Собираются и на открытом воздухе, тогда ложатся в разных направлениях, чтобы облегчить обнаружение хищников. Когда взрослые возвращаются в логово, щенки часто лижут и покусывают им морду, но срыгивания не происходит. Такое поведение переносится во взрослую жизнь.
Взрослые и молодые участвуют в играх, как правило, после отдыха или кормления. Игра может быть краткой или длиться несколько минут. Обычно игра состоит из погони, реже из борьбы.

В общении очень важны визуальные знаки и телодвижения. Важными источниками визуальных сигналов являются морда, область глаз (маска) и особенно уши и хвост. Когда лисица пристально смотрит на объект (например, на особь своего вида или шакала), голова держится высоко, глаза открыты, уши прямые и направлены вперед, рот закрыт. Когда выказывается страх или подчинение, например, при приближении хищника или другой большеухой лисицы, уши прижимаются назад, а голова держится низко. Это выражение измениться оскалом с низко опущенной головой.
Черный кончик и дорсальная полоса хвоста также являются эффективными для сигналов. Положение хвоста варьируется от висящего вниз до поднятого вверх и загнутого дугой, форме перевернутой U. Положение выгнутого хвоста очевидно при встрече с доминированием, угрозой или агрессией. Оно также используется во время сексуального возбуждения, игры и дефекации. Во время бега положение хвоста прямое горизонтальное, например, при преследовании добычи или бегстве от опасности. В экстремальных случаях угрозы шерсть на шее, плечах, крестце и хвосте может вставать дыбом, увеличивая визуальный размер лисицы. Обычно взъерошивание шерсти - реакция на приближающихся хищников и сочетается с выгнутыми спиной и хвостом.
Приветствие включает визуальные и обонятельные сигналы. Большеухие лисицы узнают особей на расстоянии до 30 м. Распознав, пристально смотрят, иногда медленно подходят или атакуют без каких-либо визуальных проявлений. Подход обычно является формой символического подчинения, в которое входят опущенная голова, вытянутая шея, прижатые назад уши и морда, направленная на угол рта другой особи. Подход подтверждается второй особью, принимающей позу с высоко поднятой головой и опущенным вниз хвостом.

Используют несколько громко издаваемых звуков. Звуки являются либо контактными знаками, либо предупреждающими и чаще встречаются зимой. Контактные звуки тихие и не слышны на большом расстоянии. Звуки предупреждения и mobbing высокие и доносятся дальше, чем контактные, но менее частые. Взрослые используют контактные звуки для подзывания щенков в логово или из него, а также для подзывания друг друга в обильную зону кормления. Запугивающие звуки используются для предупреждения других лисиц о приближении хищника.
Используют 3 позы при мочеиспускании: наклон вперед, поднятие ноги и приседание. Для физического мочеиспускания самцы обычно используют наклон вперед, а самки используют приседание. При маркировке мочой (направлении мочи на конкретный объект, обозначенный предыдущими мочеиспусканием или фекалиями) самцы используют положение с поднятой ногой, самки - приседание. Маркировка мочой происходит чаще зимой, чем летом. Иногда делаются двойные знаки, в которых первая метка самки, на которую самец помещает свою метку. Самки начинают делать метки мочой при начале эструса; ответная частота меток самцов не изменяется.
Использование выделений желез для общения неизвестно. Запахи важны во время физического контакта, который возникает, главным образом, при отдыхе вместе и во время ситуаций сближения.

Обычно игнорируют копытных. Также игнорируют белохвостых мангустов (Ichneumia albicauda), полосатых карликовых мангустов (Helogale parvula) и полосатых мангустов (Mungos mungo). Опасаются крупных хищников - львов (Panthera leo) и пятнистых гиен (Crocuta crocuta). Гиеновидные собаки (Lycaon pictus) и гепарды (Acinonyx jubatus) преследуют лисиц. Отдельные стаи гиеновидных собак специализируются на охоте на большеухих лисиц. Бурые гиены (Parahyaena brunnea), гепарды, леопарды (Panthera pardus) и львы ловят взрослых большеухих лисиц; чепрачные шакалы (Canis mesomelas) - самая большая угроза для щенков. Щенки укрываются в логовах с небольшими входами, препятствующими проникновению крупных хищников.
Группой отгоняют хищников, которые приближаются к местам размножения, включая чепрачных шакалов, стройных мангустов (Galerella sanguinea), пятнистых гиен и белохвостых мангустов. Большие пернатые хищники, такие как боевые орлы (Polemaetus bellicosus) и филины (Bubo africanus и B. lacteus), могут ловить взрослых лисиц на открытых участках. При преследовании наземными или пернатыми хищниками, большеухие лисицы быстро меняют направление, что увеличивают их шансы побега. Лисица может эффективно изменять направление при беге по плоской поверхности без потери скорости. Африканские иероглифовые питоны (Python sebae) также убивают и поедают лисиц.

Вид предрасположен и распространяет бешенство, вирус собачьей чумки и парвовирус собак. У одной большеухой лисицы из экосистемы Серенгети, Танзания, была идентифицирована Trichinella nelsoni. Вспышки бешенства с 1986 по 1989 год составляли 90% смертей среди взрослых в одной популяции лисиц. В Серенгети эпидемии бешенства являются наиболее распространенной причиной смертности во время нормального количества и плотности термитов.
Имеет коммерческое применение; местные жители Ботсваны охотятся на большеухих лисиц с апреля по июль из-за шкур. Являются эффективным и важным хищником термитов, которые считаются серьезным вредителем пастбищ.
В неволе зарегистрирована максимальная продолжительность жизни 13 лет и 9 месяцев, в дикой природе, вероятно, короче.

Литература
1. Clark H. O. Jr., 2005. "Otocyon megalotis" / MAMMALIAN SPECIES No. 766, pp. 1-5. Published 15 July 2005 by the American Society of Mammalogists
2. Nel J. A. J., Maas B. "Bat-eared fox (Otocyon megalotis)" in Sillero-Zubiri, C., Hoffmann, M. and Macdonald, D.W. (eds). 2004. Canids: Foxes, Wolves, Jackals and Dogs. Status Survey and Conservation Action Plan. IUCN/SSC Canid Specialist Group. Gland, Switzerland and Cambridge, UK. pp.183-189.